
Вы видите шоколадное печенье — последнее в коробке — и тут же хватаете его, чтобы съесть. За считанные мгновения мозг взвешивает все «за» и «против» и приводит руку в движение. Решение принимается моментально, и благодарить за это нужно электрические импульсы, с бешеной скоростью бегущие через ваш мозг. В книге «Скорость мысли. Грандиозное путешествие сквозь мозг за 2,1 секунды» (издательство «Individuum»), переведенной на русский язык Алексеем Снегировым, нейробиолог Марк Хамфрис рассказывает, что науке известно об электрических импульсах, которыми обмениваются между собой нейроны в коре головного мозга, и как с их помощью мы принимаем решения и управляем движениями тела. Публикуем фрагмент, в котором описывается путешествие импульса в первичную зрительную кору головного мозга.

Через пропасть
Наш импульс прилетает по аксону в первичную зрительную кору головного мозга, V1. Это первая из множества областей, отвечающих за обработку визуальной информации, которые составляют треть всей коры головного мозга1. Сообщение — об одном маленьком пикселе песочно-шоколадного искушения — должно пройти через все эти области, объединяясь с другими сообщениями, которые несут туда миллионы других импульсов, чтобы создать образ «печенья».
1 Некоторые из вас сейчас воскликнули про себя: «Эй, автор! А что случилось с латеральным коленчатым телом?» Импульсы не поступают напрямую из сетчатки в первичную зрительную кору. Аксоны ганглиозных клеток сетчатки приходят в латеральное коленчатое тело (ЛКТ), которое помещается на нижней стороне подушки таламуса. Один набор нейронов ЛКТ передает сигналы сетчатки в зрительные зоны коры; другие отправляют сигналы многочисленным структурам, расположенным ниже коры, таким как верхнее двухолмие среднего мозга, предназначенным для быстрой реакции на изменения в вашем поле зрения (например, уклонение от летящего в лицо футбольного мяча). Учитывая тридцать каналов, исходящих из сетчатки, существует, как вы можете себе представить, не менее сложный набор каналов, выходящих из ЛКТ, которые пока только исследуются.
Но сначала нам надо добраться до первого пункта назначения. Кора головного мозга представляет собой что-то вроде торта из шести слоев; начинка, состоящая из нейронов, есть во всех слоях кроме первого, верхнего. Вместе с нашим импульсом мы собираемся достичь конца аксона в четвертом слое зрительной зоны V1. Над нами — три этажа нейронов второго и третьего слоя; а еще выше — первый слой, где изредка можно встретить маленький нейрон, но в основном он состоит из аксонов, идущих в другие слои и места, и вспомогательных клеток, которые не являются нейронами, — клеток глии, которые выполняют функции строительных лесов, уборщиков и чернорабочих. Под нами — большие, можно даже сказать гигантские клетки нейронов пятого и шестого слоев.
Тела нейронов могут быть упакованы слоями, но бóльшая их часть располагается довольно свободно. Мы оказываемся в своеобразном лесу. Из тела каждого нейрона вырастает огромное дерево. Тонкие ветви раздваиваются, разветвляются, искривляются. Они занимают гораздо больше места, чем тела нейронов. Эти деревья — дендриты, отростки, через которые нейрон получает входные данные, импульсы, от других нейронов и отправляет их к своему телу. Аксоны от бесчисленного количества других нейронов упираются в кончики веток дендритных деревьев окружающих нас нейронов.
Форма дендритов и их количество может многое сказать о том, для чего предназначен нейрон. Действительно, исторически именно по их количеству и расположению мы часто отличали нейроны друг от друга. Наше путешествие с импульсом от сетчатки вот-вот приведет нас к компактному дереву дендритов первого — на нашем пути — звездчатого нейрона в коре головного мозга. Под нами находится типичный нейрон коры, пирамидальная клетка пятого слоя, обладающая двумя видами дендритных деревьев: одно выходит из верхушки конуса, длинный тонкий стебель тянется почти до кортикальной поверхности; другое, густое и раскидистое, расположено ниже тела клетки (рис. 3.1). Над нами, во втором и третьем слоях, — более скромные пирамидальные нейроны, их деревья компактны и окружают тело, не так привлекая внимание, как их старшие братья в пятом слое. Независимо от формы и размера2 все эти дендриты подключены к сигнальным кабелям, идущим от других нейронов.
2 Нейроны бывают самых разных форм. Множество примеров форм кортикальных нейронов, а также многих других типов нервных клеток можно посмотреть на http://NeuroMorpho.org.
Но как только мы погружаемся в четвертый слой и достигаем конца аксона, бег нашего импульса резко останавливается, упершись в преграду. Между концом аксона и началом дендрита следующего нейрона есть промежуток — синаптическая щель, — который непреодолим для электрического импульса. Как передать послание, которое несет импульс? Как пересечь эту пропасть и создать новый импульс в следующем нейроне, чтобы восстановить сообщение?
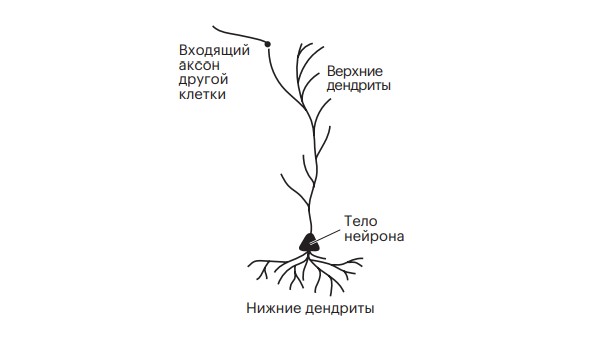
Рисунок 3.1. Пирамидальный нейрон пятого слоя коры. Он называется так потому, что его тело имеет форму трехмерной пирамиды.
Прибытие нашего электрического импульса вскрывает пузырьки с молекулами, хранящимися на конце аксона, заставляя их содержимое выделяться в промежуток между нервными окончаниями, а химическая диффузия отправляет их на другую сторону промежутка (рис. 3.2). Когда эти молекулы захватываются дендритом принимающего нейрона, они немного изменяют его электрический потенциал. Но совсем чуть-чуть. Причем этот небольшой скачок напряжения происходит на самом дальнем конце довольно длинного дендрита следующего нейрона, вдали от тела клетки. Какое именно воздействие окажет прибывший по аксону импульс на дендрит, зависит от того, молекулы какого вещества он пошлет через промежуток между ними. У нейронов одного типа пузырьки на конце аксона содержат молекулы одного и того же химического вещества-нейромедиатора. Но разные типы нейронов могут отправлять в промежуток разные нейромедиаторы, а от типа молекулы зависит, будет скачок потенциала в принимающем нейроне-мишени вверх или вниз.
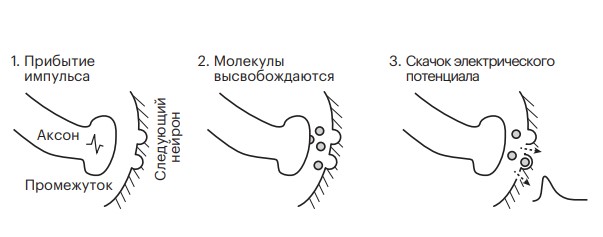
Рисунок 3.2. Отправка импульса через промежуток между нейронами.
Наш импульс, несущий информацию о печенье, стремительно влетев в терминальный конец аксона, «лопает» пузырьки — разрывает «упаковки» с молекулами глутамата. Освободившись, молекулы проходят через мембрану, диффундируют в растворе, заполняющем промежуток шириной в микрометр, и натыкаются на рецепторы глутамата с другой стороны мембраны дендрита. Если молекула прибудет туда в правильной ориентации, она плотно зафиксируется на рецепторе — процесс напоминает игру двухлетнего ребенка с кусочками пазла, когда, случайным образом смешивая их вместе, у него иногда получается вставить торчащий выступ одного кусочка в вырез другого. Захват рецепторами молекул вызывает открытие расположенных вокруг них каналов в мембране нейрона. Ионы, устремившиеся через каналы внутрь клетки, создают скачок напряжения на этом участке дендритного дерева, принимающего сигнал нейрона. Этот рецептор ищет именно глутамат, поэтому поток ионов создает на целевом нейроне небольшое увеличение электрического потенциала, положительный импульс. Мы называем это возбуждением.
Рядом, немного дальше по дереву дендритов, ближе к телу принимающего нейрона, мы видим терминалы других аксонов, которые приходят туда не из сетчатки. Скорее всего, они принадлежат небольшим нейронам, изредка встречающимся поблизости тут и там. И они, по прибытии в терминальный конец импульса, будут отправлять через промежуток другое вещество, молекулу γ-аминомасляной кислоты (ГАМК). Когда ГАМК поступает на соответствующие рецепторы на том же самом дендритном дереве, она вызывает поток ионов, уменьшающих потенциал клетки в этом месте, и создает отрицательный импульс. Следуя логике, мы называем это торможением.
Когда после взаимодействия между терминальным концом аксона и принимающим концом дендрита целевого нейрона в нем происходит этот скачок потенциала — напряжение повышается или понижается, — этот импульс распространяется по дендритному дереву, от синаптического промежутка до тела целевого нейрона. Слабея по мере продвижения, электрический всплеск немного увеличивает или уменьшает постоянно изменяющийся электрический потенциал на теле принимающего нейрона, суммируясь с другими. И, возможно, способствует возникновению в нем нового импульса: пришедший положительный импульс возбуждения заставит целевой нейрон с большей вероятностью создать новый импульс, отрицательный импульс торможения понизит эту вероятность.
Процесс передачи импульса принимающему нейрону кажется немного безумным. Ваш мозг приложил огромные усилия, чтобы создать импульс — процесс весьма энергозатратный, — чтобы обойти ограничения отправки сообщений на большие расстояния путем простого выброса химических веществ или распространения скачков электрического потенциала. И все лишь для того, чтобы затем снова превратить импульс в выброс молекул нейромедиатора, которые вызывают скачки электрического потенциала клетки.
Но тому есть веские причины. Например, передача сигнала с помощью химии и потенциала намного менее энергозатратна — в организмах с крошечным мозгом все сигналы передаются за счет распространения скачков потенциала и химических веществ, а не с помощью импульсов. Но, возможно, основная причина — гибкость. Преобразование импульсов обратно в химическую, а затем электрическую форму дает мозгу возможность по-разному оперировать с одним и тем же бинарным («все или ничего») импульсом.
Гибкость возникает из-за разницы в строении синаптических щелей. Синапсы одного и того же типа, которые содержат пузырьки с одинаковыми нейромедиаторами, необязательно производят всплеск электрического потенциала одинаковой величины. Эта разница в амплитуде может объясняться некоторыми изменениями по обе стороны от синаптической щели. Например, нейрон-мишень может обладать бóльшим количеством рецепторов, принимающих молекулы нейромедиатора: чем больше рецепторов будет заблокировано, тем больше ионных каналов откроется в мембране и тем больше будет скачок потенциала. Проницательный читатель, вероятно, уже догадался, что мы также можем увеличить амплитуду напряжения, выбрасывая больше молекул нейромедиатора в щель — тем большее число молекул случайно окажется в правильном положении в нужном месте и заблокирует большее количество рецепторов. Все это означает, что приходящий импульс может быть преобразован из бинарного сигнала «все или ничего» в аналоговый всплеск напряжения с некоторым коэффициентом, определяющим диапазон воздействий на нейрон-мишень.
Но есть строгие ограничения на размеры одного синаптического промежутка. Вся конструкция — терминальный конец аксона, синаптическая щель, рецепторы на другой стороне — составляет лишь несколько микрометров в поперечнике. В этом пространстве может разместиться только определенное количество рецепторов, а терминал аксона может хранить только ограниченное количество пузырьков с молекулами нейромедиатора. Эти жесткие рамки означают, что одного приходящего импульса недостаточно для создания нового импульса. Именно поэтому мы прибыли с одним из миллиона импульсов, отправленных сетчаткой: нам нужна целая армия импульсов, чтобы сгенерировать новый.
Подробнее читайте:
Хамфрис, М. Скорость мысли. Грандиозное путешествие сквозь мозг за 2,1 секунды / Марк Хамфрис ; [пер. с английского Алексея Снигирова]. — Москва: Individuum, 2022. — 304 с.