
Больше трех миллиардов лет эволюции животные провели без нервной системы — по крайней мере в том виде, как мы ее знаем. Да и сейчас значительное количество организмов легко выживают без нервных клеток и мозга. Появление нейронов — как будто маловероятная загадочная случайность, о которой по-прежнему вопросов больше, чем ответов. Какими были первые нервные системы? Что унаследовал современный мозг от своих древних предков? Что было раньше: нейрон или нейротрансмиттер?

По нейросекреторной гипотезе, то, чем мы задумываемся над этими вопросами, — всего лишь побочный продукт эволюции. Ничто не требовало появления нейронов у древних многоклеточных организмов. Зато у предков многоклеточных были ферменты, маленькие пептиды, рецепторы, токсины — площадка для молекулярных игр, на которой по счастливой случайности собралась нервная система.
Эта гипотеза происхождения нервной системы подползла к правде ближе остальных. Она предполагает, что в прошлом существовали какие-то донервные сигнальные системы. Некоторые из этих систем исчезли, некоторые — сохранились и у современных животных. И если мы хотим лучше понять, как появилось то, что в голове у нас, стоит пристальнее посмотреть на тех, у кого даже головы нет. Оказалось, что трихоплакс — примитивный живой блинчик размером около двух миллиметров — очень похож на маленький мозг. Но как у довольно неразумной пластинки появились аналоги нейронов и синапсов? И что эти аналоги могут рассказать об эволюции нервной системы?
Раз — ступенька, два — ступенька
В школе про эволюцию нервной системы рассказывают сдержанно.
Вот амеба: одноклеточное, у которого нет и не может быть нейронов. Но хотя думать ей вроде как нечем, она реагирует на раздражители из внешней среды. Например у амебы есть отрицательный таксис: она уползает оттуда, где ей плохо, — подальше от света. Вот гидра. У гидры с нейронами получше, чем у амебы, но организации в них немного: они разбросаны тут и там по ее телу. Если ткнуть гидру палкой, все ее тело сожмется. Так работает диффузная нервная система. В нейронах плоских и круглых червей появляется больше порядка: они собираются в нервные стволы. Стволы соединены друг с другом перемычками, словно две веревки соединены ступенями веревочной лестницы. Ступенька за ступенькой собралась нервная система лестничного типа.
У кольчатых червей в той части тела, которая больше всего похожа на голову, нейроны сбиваются в кучки, надглоточный и подглоточный ганглий, а нервные стволы сливаются в единую брюшную цепочку. У первых хордовых, наших далеких предков, случается хитрый разворот: нервная трубка идет не вдоль брюха, а вдоль спины, и даром что головного мозга нет. У рыб, наконец, появляется мозг, а к млекопитающим и птицам он достигает небывалой сложности.
Логика этой истории — прямая и бесхитростная, как нервная цепочка у червяка. Но как все бесхитростное, она крошится и осыпается, стоит только поскрести.
Стройная последовательная система рушится, даже если просто посмотреть на эволюционное дерево. Все ломается в самом начале, когда оказывается, что развитие нервной системы — никак не может быть прямой линией, это точно несколько изолированных друг от друга, ветвящихся эволюционных лучей. Гидры, например, относятся к совсем другому таксону многоклеточных — стрекающим (Cnidaria), в то время как черви, пчелы, рыбы и мы — двусторонне-симметричные (Bilateria). В школьной истории к тому же напрочь упущен еще один базовый таксон многоклеточных животных с нервной системой — гребневики (Ctenophora).
Долгое время считалось, что нейроны стрекающих, двусторонне-симметричных и гребневиков имеют единое происхождение. Усомниться в этом удалось лишь недавно, в 2010-х, когда набрали силу молекулярные методы анализа генома. Даже первоначальные данные показали, что нейроны и центральная нервная система, весьма вероятно, появились более одного раза у различных эволюционных линий. У одних только билатерий развитие головного отдела нервной системы могло происходить независимо по меньшей мере пять раз: у хордовых, членистоногих, нематод, кольчатых червей и моллюсков.
Нервная система книдарий тоже оказалась вовсе не так проста, чтобы стоять так близко к началу эволюционной цепочки. Гидры вытягиваются, покачивают щупальцами, кувыркаются, сжимаются и используют щупальца для кормления. Кувыркание и кормление — довольно сложные формы поведения, которые требуют последовательно выполнить серию движений, а значит — переключаться между несколькими двигательными программами в нервной системе. Регистрация активности нейронов гидры показала, что при разных формах поведения активируются различные нейронные сети. Нейроны книдарий отличаются от нейронов двусторонне-симметричных: в их синапсах сигнал может проходить в обе стороны, в то время как у билатерий синапсы допускают только одностороннее движение.
Гребневиков же долго относили в одну группу с книдариями — Coelenterata. Но геномный анализ сначала позволил отделить гребневиков в обособленный таксон, а затем показал, что они заметно отличаются от других животных по нейронным генам: гены «классических» нейромедиаторов и рецепторов к ним либо не экспрессируются в нейронах, либо вообще отсутствуют. Кроме того, у гребневиков две нервных системы: одна расположена в мезоглее (это сильно обводненная соединительная ткань, расположенная между двумя эпителиями) другая — в эктодерме (это внешняя эпителиальная ткань). Нейроны мезоглеи и эктодермы отличаются друг от друга по набору белков и особенностям формирования сети.
Так что нервные системы гребневиков, книдарий и двусторонне-симметричных, скорее всего, возникли изолированно на давно разошедшихся эволюционных ветвях. Поэтому и нейроны у них получились удивительно непохожими друг на друга.
Впрочем, чтобы заметить их непохожесть, ученым пришлось для начала заметить, что нейроны — обособленные клетки, а не цельные проводки. Автор «нейронной доктрины» испанский гистолог Сантьяго Рамон-и-Кахаль в конце XIX века обратил внимание, что отдельные нервные клетки и отростки хоть и прилежат очень тесно друг к другу, но все-таки не сливаются; и между двумя утолщениями отростков этих клеток остается крохотная щель — синапс, через который проходит импульс. То, как через синапс проходит сигнал, понятно тоже было не сразу. Нейрофизиологи оставались приверженцами электрической теории, в то время как фармакологи настаивали, что нейроны взаимодействуют, выделяя химические вещества. В этой войне супов и искр победили супы: первое поражение искрам нанесло открытие нейротрансмиттеров ацетилхолина и норадреналина, а окончательным нокаутом стало обнаружение мембранных пузырьков с нейротрансмиттером в нервных окончаниях с помощью электронной микроскопии.
Зачем нужны пузырьки? ↓
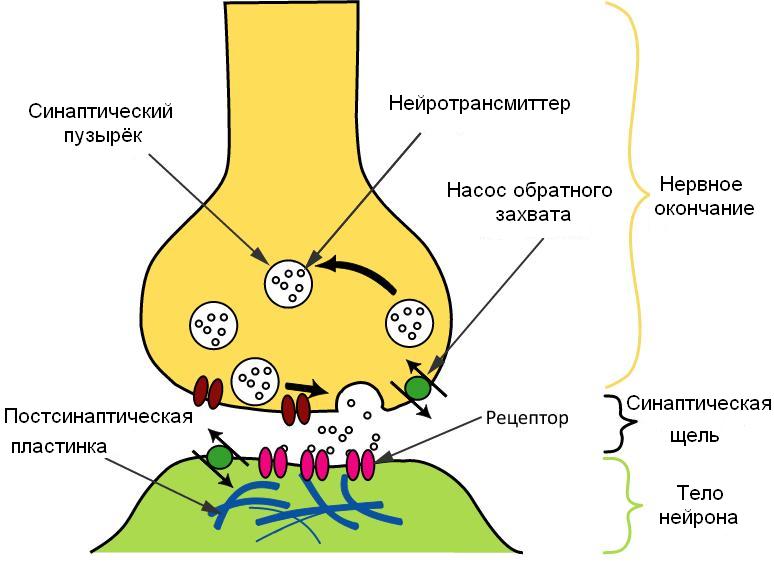
Но к концу ХХ века вдобавок к ацетилхолину и норадреналину отыскали еще несколько десятков нейромедиаторов и тысячи нейропептидов. И стало совершенно неочевидно, зачем такое обилие, когда можно было обойтись двумя: возбуждающим и тормозящим. Или даже одним медиатором, при условии, что к нему существовали бы два типа рецепторов — возбуждающие и тормозящие.
Тем более непонятно, как такая избыточная система возникла трижды у далеко отстоящих друг от друга многоклеточных.
Шиворот-навыворот
В середине XX века, когда электрическая парадигма нервной системы уступила химической, ученые предположили, что раз уж нервные клетки регулярно выделяют медиаторы вовне, должно быть, они произошли из секреторных клеток. К тому же ацетилхолин, норадреналин и другие медиаторы обнаружились у донервных организмов, в том числе у одноклеточных инфузорий, которые в принципе не могли обладать нейронами. В результате возникла нейросекреторная теория эволюции нервной системы. А многообразие нейротрансмиттеров и нервных клеток у современных животных большинство ученых поначалу объясняло дивергенцией — постепенным расхождением признаков какой-то одной предковой клетки.
Но в 1974 Тот Дмитрий Сахаров, который под псевдонимом Дмитрий Сухарев написал песню про пчелу, жужжащую про Чимган и Бричмуллу.
Объемная передача медлительна, но в протерозойском мире без крупных быстрых хищников ранним животным этой скорости было достаточно для координации поведения.
Однако такой тип передачи сигналов в нервной системе существует и сейчас. Например, одна из простейших сигнальных молекул мозга, оксид азота (II), выделяясь из нейрона, действует на множество клеток, не находящихся в непосредственном контакте с нейроном-источником NO. Так же и дофамин в стриатуме выделяется через «открытые» синапсы — то есть из аксонов дофамин выбрасывается не в сторону соседней клетки, а в окружающую межклеточную жидкость.
Иногда, выплескиваясь мимо синапсов, нейротрансмиттеры размывают границу между нервным и гормональным. Ведь медиаторы работают не только в нервной системе, а приставка нейро- зачастую весьма условна. Многие нейроны одним окончанием формируют обычный синапс, а другим — высвобождают ту же молекулу в кровоток или другие циркуляторные системы в качестве гормона. Например, у медицинских пиявок клетки Ретциуса при низкочастотной активации ведут себя как типичные нейроны, но при высокочастотной активации выбрасывают серотонин уже как гормон, контролирующий питание и выделение слизи.
Нейрофизиологи считают объемную передачу и гормональные функции нейронов неканоническими, потому что их открыли после изучения передачи сигналов в нервной системе. Кроме того, неканоническим является и воздействие нейромедиаторов на ядерные белки (гистоны) или выделение АТФ и глутамата через белки Электрические синапсы — синапсы, в которых происходит прямая передача сигнала от нейрона к нейрону через специализированный щелевой контакт без выделения медиатора.
Медиаторы работали как гормоны и передавали сигналы объемно задолго до появления нейронов и синапсов. Трансмиттеры объединили разнородные популяции клеток в системы управления поведением донервных организмов. Они стали трансмиттерами до того, как приросли приставкой нейро-. Не нервная система создала трансмиттеры, наоборот, трансмиттеры создали нервную систему.
Синапсы появились только когда потребовалась более быстрая, а значит — и более локальная передача сигнала. Возможно, сыграло роль и то, что выделение медиаторов в замкнутых объемах требовало меньше ресурсов и было энергетически выгоднее, чем выделение большого количества молекул в ничем не ограниченный объем. Наконец, так как некоторые из медиаторов, например, глутамат, могут быть источниками пищи — попытка запереть их в синапсы могла быть связана с необходимостью защитить их от поедания другими клетками, симбионтами или паразитами.
Но если нервная система появилась из-за трансмиттеров, то как появились сами трансмиттеры?
В начале была травма
Объяснить, почему возникло такое разнообразное множество медиаторов, можно с двух точек зрения — эволюционной и функциональной. Но это не противоречащие друг другу альтернативные сценарии, а скорее взаимодополняющие пути создания нервной системы.
Эволюционный сценарий предполагает, что разные типы нейронов — наследство из разнообразных секреторных клеток от ранних животных. Второй, функциональный, сценарий добавляет, что молекулы-передатчики выполняли и продолжают выполнять разные функции в клетках, синапсах и за их пределами. Медиаторы могут организовывать различные формы поведения: серотонин у пиявок стимулирует пищевое поведение, у жужжащих пчел нейромедиатор октопамин вовлечен в развитие агрессии, у морских зайцев (аплизий) нейрогормоны и нейромедиаторы способствуют размножению. Скорее всего, у ранних донервных животных медиаторы действовали как интеграторы поведения. При этом большое количество возможных сценариев поведения для своей координации требовало и большого количества медиаторов и их сочетаний.
Общая гипотеза заключается в том, что сигнальные молекулы первоначально возникли для объемной передачи самых разных сигналов: они были и иммунными сигналами, и гормональными, и донервными, и пищевыми. Уже потом это функциональное множество было приспособлено именно в качестве передатчиков между нейронами.
Одними из трансмиттеров в этом множестве, вероятно, стали молекулы, выделяющиеся при повреждении: АТФ, NO, протоны и глутамат. Глутамат — один из основных нейромедиаторов нервной системы, и упрощенно говоря, мозг — это глутаматергическая машина. Но в то же время многие организмы (включая растения) используют глутамат как сигнальную молекулу в случае повреждений. Оксид азота — тоже одна из самых универсальных сигнальных молекул, которая связана с реакцией на травму у всего живого — от растений до архей. Но при этом нитрергические нейроны составляют всего около двух процентов популяций клеток в мозге, где NO действует как классический нейромедиатор.
Спусковым крючком для развития нейромедиаторов могло стать и возникновение внеклеточного пищеварения. Чтобы переварить еду снаружи, древним организмам приходилось выделять большое количество молекул, которые затем стали нейротрансмиттерами. К тому же пищу было необходимо очистить от патогенов, вроде бактерий, чтобы защитить себя.
Первые иммунные молекулы тоже могли в конечном счете стать предковой формой трансмиттеров. Тот же оксид азота — один из важнейших медиаторов воспаления и врожденного иммунного ответа.
При этом у далекого предка многоклеточных границу между пра-пра-иммунным, пра-пра-гормональным и пра-пра-нервным было не разглядеть. Только позже в ходе эволюции эти молекулы стали частью в меру изолированных сигнальных систем: нервной, гормональной и иммунной. Хотя и сейчас граница между ними размытая, и сигнальные системы продолжают функционально дублировать друг друга. Гипоталамо-гипофизарная система мозга, например, выполняет и функции нервной системы, и гормональной. Нейроны задней доли гипофиза выделяют в кровоток гормоны окситоцин и вазопрессин, которые влияют на водный обмен и социальное поведение. Нейроны передней доли воздействуют нейрогормонами (либеринами и статинам) и на передний гипофиз. А гипофиз уже выделяет гормоны, действующие на периферические органы.
Все это было бы простыми предположениями и рассуждениями о прошлом, если бы в спокойных тропических водах не жили организмы, которые оказались удивительно похожим на животное с донервной сигнальной системой, каким его представлял Сахаров.
Ползучее подтверждение
Пластинчатые — небольшие многоклеточные существа, которых считают самыми просто устроенными животными. Изначально тип Placozoa учредили лишь для единственного вида — трихоплакса (Trichoplax adhaerens). Сейчас у него нашли еще двух близких родственников: Hoilungia hongkongensis и Polyplacotoma mediterranea. Выглядят плакозои как маленькие блинчики размером от 1 до 3 миллиметров и толщиной в пару клеток. У тела трихоплакса нет ни переда и зада, ни права и лева.
Тело пластинчатых — трехслойный пирог. Нижний слой состоит из эпителиальных клеток, густо усеянных длинными ресничками для передвижения. Между эпителиальными клетками находятся железистые и липофильные клетки для пищеварения. Верхний эпителиальный слой не может похвастаться густой шевелюрой из ресничек, его клетки сплющены, а реснички — редкие и короткие. «Спинка» защищает трихоплакса от хищников. Между верхним и нижним слоем, в сердцевине, находятся звездчатые клетки, которые почти сливаются в единый синцитий, как мицелий грибов, клетки с капельками липидов, и клетки с кристаллами карбоната кальция. По-видимому, эти кристаллические клетки помогают пластинчатым чувствовать изменение положения своего скромного тела в пространстве.
Ни нейронов, ни чего-то, напоминающего нейроны, среди клеток трихоплакса нет. Тем не менее эти животные целенаправленно ползут к еде и свету. Трихоплакс может зависнуть над пищей, а переварив ее — двинуться дальше. Выяснилось даже, что к перевариванию водорослей безумный трихоплакс подходит с умом.
При контакте с водорослями липофильные клетки выбрасывают содержимое максимально рационально: в переваривании пищи участвуют только клетки поблизости от скопления пищи, причем если водорослей мало — выделяют гранулы лишь некоторые клетки, а если пищи больше — то и больше клеток секретируют гранулы. После переваривания пищи трихоплакс взбалтывает то, что переварил: тело трихоплакса выгибается вверх, клетки по краю тела остаются неподвижными, а клетки в центре двигаются по эллиптическим траекториям. Возможно взбалтывание помогает быстрее впитывать питательные вещества. Кроме того, в каком-то смысле трихоплаксы роятся — собираются плоской толпой из нескольких организмов, чтобы съесть водоросль вместе. Такое роение считают формой социального поведения.
Любая из этих форм поведения требует сложной координации клеток. Ведь при беспорядочном биении ресничек эпителия животное просто вертелось бы на месте. И если до обнаружения такого сложного поведения трихоплакс был всего-навсего одним из нескольких сотен примитивных животных, которым в сборниках уделяют десяток строчек, то теперь он немедленно перешел в ранг животных, достойных самого пристального внимания. Как же договариваются клетки животного, не отягощенного ни нервной системой, ни нейронами, ни синапсами?
Последователи Сахарова, Дарья Романова и Леонид Мороз, обнаружили, что координацию обеспечивают широко распространенные медиаторы глутамат, АТФ, гамма-аминомасляная кислота (ГАМК) и глицин. Глутамат вызывает циклы кормления, а ГАМК — подавляет. Ученые доказали, что эти вещества не появляются из внешней среды, а их выделяют клетки трихоплакса.
Чтобы показать это, биологи поместили пластинку в среду без кальция. Кальций необходим для выделения пузырьков с молекулами во внешнюю среду, без него любые сигнальные молекулы застрянут в клетке. А если эти молекулы отвечают за скоординированное движение трихоплакса, то без кальция в среде трихоплакс растеряется и будет вертеться на месте. Именно так себя вели пластинчатые в среде без кальция: каждая ресничка и клетка нижнего эпителия утрачивала способность общаться с окружением и двигалась сама по себе. В результате трихоплакс хаотично топтался на месте, а не полз куда-то что-то есть.
У трихоплакса нашли и гены компонентов синаптических пузырьков и регуляторов их высвобождения.
Но для объемной передачи мало выделить сигнальные молекулы. Чтобы цепочка сработала, у других клеток трихоплакса должны быть рецепторы к этим веществам. В геноме трихоплакса закодированы рецепторы к АТФ, ГАМК и глутамату. А рецептором для глицина может быть та же структура, которая отвечает за восприятие глутамата. При этом репертуар рецепторов у пластинчатых богаче, чем у любого другого многоклеточного, включая млекопитающих и человека, — 85 типов рецепторов. В генах пластинчатых нашли и другое важное для нервной системы звено — закодированные ионные каналы. У животных с нервной системой ионные каналы участвуют в формировании необходимого для распространения нервного импульса потенциала действия.
У двух представителей пластинчатых, Trichoplax и Hoilungia, в геноме представлены по крайней мере пять разновидностей ионных каналов, а у представителей второго — семь. Так что у непростого трихоплакса с одной стороны налажено выделение сигнальных молекул, а с другой — есть все необходимое, чтобы воспринять сигнал и передать его дальше. Подобно крохотной планете-океану Солярис, которая, как оказалось, сознательно меняет свою орбиту, трихоплакс, казалось бы простой и примитивный, целенаправленно куда-то ползет и что-то ест. И делает это благодаря всем тем же медиаторам и рецепторам, которые используют люди, его изучающие.
***
Трихоплакс — возможно, самое близкое подобие Urmetazoa, гипотетического предка многоклеточных, поэтому он позволяет посмотреть на первые шаги эволюции нервной системы. Чтобы координировать довольно сложное поведение, этой пластинке не нужны ни синапсы, ни нейроны, — то есть первые сигнальные системы могли обходиться без специализированных клеток, с помощью объемной передачи молекул. Без чего они не могли обойтись — так это основных компонентов системы по отдельности: трансмиттеров, рецепторов и ионных каналов. Все эти компоненты обнаружили у трихоплакса, некоторые — в большем разнообразии, чем у других многоклеточных.
По дороге к современным нервным системам часть этого биохимического обилия была утрачена, часть стала основой других регуляторных систем, иммунной и гормональной, а часть — специализировалась именно на передаче химико-электрических импульсов в нервной системе. В угоду себе трансмиттеры превратили секреторные клетки в нейроны, а объемную передачу — в куда более быструю и эффективную передачу в синапсах.
Нейросекреторная гипотеза, которую, казалось бы, так убедительно доказывает трихоплакс, не проясняет, в какой момент появились специализированные секреторные клетки. Может ли трихоплакс рассказать и про это? И хотя понятно, что он использует нейромедиаторы в качестве сигнальных молекул для объемной передачи, пока совершенно непонятно, откуда они берутся. Непонятно, есть ли у пластинчатых специфические пептидергические, глутаматергические или ГАМКергические клетки. И если есть — где они находятся на этом плоском маленьком теле? Возможно, самые первые источники медиаторов были не эндогенными, а экзогенными. То есть клетки не синтезировали глутамат или ГАМК, а поглощали их из внеклеточной среды, задерживали в себе и в нужный момент — выбрасывали обратно. Оба сценария возможны, но найдется ли доказательство одного из них у близкого подобия Urmetazoa?
Чтобы гипотеза стала теорией, ученым предстоит еще долго присматриваться к трихоплаксу. И надеяться, что трихоплакс не начнет присматриваться в ответ.